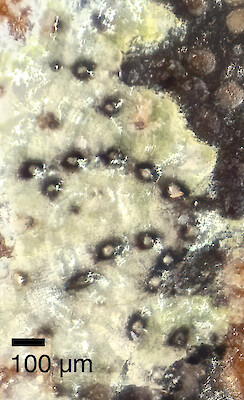
Strigula oleistrata






Common names
pizza lichen
Biostatus
Native – Endemic taxon
Interim conservation status
Please note, conservation statuses are often proposed by authors of newly described taxa when they are published between NZTCS assessment periods. If such proposed conservation statuses are consistent with the criteria of the NZTCS, they are presented here as ‘interim’ statuses in accordance with the NZTCS manual, whilst acknowledging that they have not been formally assessed by the NZTCS panel.
2019 | At Risk – Naturally Uncommon | Qualifiers: DP, RR, Sp. | Interim | Ford M., Blanchon D.J., Veale A., Doyle E.J., Rolfe J.R., and de Lange P.J. (2019)
Category
Lichen
Structural class
Lichens - Crustose
Lichen substrate
Foliicolous (leaves)
Substrate details
Foliicolous
Threats
Sites where this species has been confirmed from appear to have both S. oleistrata and S. novae-zelandiae in similar abundances. Under these circumstances a conservation assessment comparable to that of S. novae-zelandiae is justified. Therefore, Strigula oleistrata has been provisionally assesed as ‘At Risk-Naturally Uncommon’ (Ford et al. 2019).
Detailed taxonomy
Genus
Family
Endemic taxon
Yes
Endemic genus
No
Endemic family
No
Other information
Etymology
oleistrata: Meaning ‘strewn with olives’
Referencing and citations
References and further reading
Ford M., Blanchon D.J., Veale A., Doyle E.J., Rolfe J.R., and de Lange P.J. 2019: Hidden in plain sight—a new Strigula species segregated from Strigula novae-zelandiae (Lichenized Ascomycota: Strigulaceae). Phytotaxa, 424(5): 267-281.
Sérusiaux E. 1998: Further observations on the lichen genus Strigula in New Zealand. The Bryologist 101: 147–152. https://doi.org/10.1639/0007-2745(1998)101[147:FOOTLG]2.0.CO;2
Jiang S.H., Wei X.L. and Wei J.C. 2016: Strigula sinoaustralis sp. nov. and three Strigula spp. new to China. Mycotaxon 131: 795‒803. https://doi.org/10.5248/131.795
Jiang S.H., Wei X.L. and Wei J.C. 2017: Two new species of Strigula (lichenised Dothideomycetes, Ascomycota) from China, with a key to the Chinese foliicolous species. Mycotaxa 19: 31‒42